את ספיגת החמצן מן הסביבה ופליטת הפחמן הדו-חמצני מהגוף מכנים מדענים, מחברי ספרים ומורים "חילוף הגזים של הנשימה". מזל שדגים אינם לומדים ואינם מרבים לקרוא ספרים
מאז הפכו היצורים הפוטוסינתטיים את אטמוספרת כדור-הארץ למאגר עצום של חמצן מולקולרי (O2), עומד לרשותם של היצורים המכונים "אווירניים" (אאירוביים) מקור עשיר מאד של אנרגיה כימית הנדרשת לפעולות החיים – המערכת: חומרים אורגניים - חמצן מולקולרי. חימצון של חומרים אורגניים בעזרת חמצן מולקולרי הוא תהליך יעיל מאד לשם זירחון של ADP (אדנוזין דו-זרחתי) והפיכתו ל-ATP, חומר "עתיר אנרגיה כימית".
ה-ATP הוא חומר מזרחן, המניע תהליכים חיוניים רבים בתאים חיים, כגון בניית חלבונים, העברת יונים בעזרת "משאבות יונים" והפעלת חלבונים מתכווצים המאפשרים תנועה. חימצון של חומרים אורגניים בעזרת חמצן מולקולרי שבאוויר מאפשר גם פעולה של מנועי השריפה במכוניות ובמטוסים. הפקת חשמל בתחנות כוח השורפות סולר, בישול באמצעות כיריים הפועלים על גז הבישול, יצירת אור באמצעות מנורות שמן ונרות פרפין, והוא גם אחראי – כך מתברר (ראו: אבי בורג ויהושע קולודני – "סלעים בוערים בישראל", גליליאו 61) – ליצירתם של סלעים מטמורפיים הנמנים על "תצורת חתרורים", וזאת בעקבות שריפת החומר האורגני ביטומן שהכילו הסלעים.
ביצורים אאוקריוטים (בעלי גרעין-תא מוקף בקרומית; הכוונה לבעלי-חיים ולפרוטוזואונים, לצמחים ולפטריות; בניגוד לחיידקים) החמצן המולקולרי מחמצֵן חומרים אורגניים תוך "רתימת" התהליך לחידוש מלאי ה-ATP באברוני תא זעירים המכונים מיטוכונדריונים. המיטוכונדריון הוא אברון זעיר, אך מורכב מאד מבחינה ביוכימית ומִבנית – פגיעה בשלמותו פוגעת בצימוד בין חימצונם של חומרים אורגניים לבין בניית ה-ATP.
 |
קטע זעיר מריאה של אדם. בחלל הנאדית נמצאים חמצן ופחמן דו-חמצני (כמו גם חנקן, גזים אצילים ואדי מים) בפאזה גזית, אך בנימי הדם אין פאזה גזית, אלא תמיסה מימית ובה גופיפי דם.
מכל מקום, חמצן מולקולרי הוא בבחינת "אוויר לנשימה" עבורנו רק עקב השתתפותו בתהליך חימצון שמתרחש במיטוכונדריונים; תנועות הנשימה כשלעצמן אינן אלא אמצעי להעברה יעילה של חמצן מן האוויר אל הדם והעברת פחמן דו-חמצני מהדם אל האוויר. למרות שבמהלך ההיסטוריה האנושית מקובל היה לראות בתנועות הנשימה, שאיפת האוויר והנשיפה, איזו מהות הצופנת בחובה את עצם החיים (כפי שאפשר להיווכח מביטויים כגון "נשמת אפו" או "רוח חיים"), הרי שבעקבות פיתוחה של מכונה המסוגלת להוסיף חמצן ישירות לדם ולסלק ממנו CO2 ניתן להחזיק אדם בחיים מבלי שיעשה כלל תנועות שאיפה ונשיפה. מכונות כאלו, מכונות "לב-ריאות", קיימות מזה כחמישים שנה, והן מאפשרות, בין השאר, עריכת ניתוחי לב פתוחים.
ברבים מבעלי החיים יש מערכות זרימה מיוחדות שתפקידן לשנע חמצן מולקולרי שנספג מן הסביבה החיצונית אל תאי הגוף; בחולייתנים עושה זאת הדם המוביל חמצן מולקולרי (O2) שנספג מן הסביבה החיצונית אל רקמות הגוף השונות. האיברים בהם נעשית ספיגה של חמצן מן הסביבה אל הדם מכונים איברי נשימה – בחולייתנים אלה הם הזימים, העור החיצוני (בעיקר בדו-חיים) והריאות.
בדם החולייתנים כלול חלבון מורכב מיוחד, המוגלובין, המגדיל מאד את כושר נשיאת החמצן מאיברי הנשימה לרקמות הגוף; שהרי מסיסות החמצן בפלזמת הדם (הנוזל שבו שרויים תאי הדם) קטנה למדי. החרקים עושים שימוש ב"פטנט" מיוחד להם – צינורות נשימה (טרכֵיאות) המובילים אוויר הישר לרקמות; גוף החרק מפולש בהם.
מן האוויר אל התמיסה
איברי הנשימה של הזוחלים, העופות והיונקים – וכן של צפרדעים וקרפדות בוגרות – הם ריאות (בדו-חיים גם לעור יש תרומה משמעותית). ריאות הן שקעים פנימה אל הגוף, שקעים שבדופנותיהם נימי דם סמוכים מאד לחללי אוויר שמקושרים אל האוויר החיצוני באמצעות מערכת צינורות – דרכי הנשימה. חמצן מולקולרי עובר מן האוויר אל הדם שבנימים, ומובל עם זרם הדם לרקמות הגוף.
יעילות העברת החמצן מהאוויר לנימי הדם מוגברת על ידי הקיפולים הרבים – נאדיות הריאה. ריאותיו של אדם כוללות כ-300 מיליון נאדיות, שלהן שטח כולל של כ-70 מ"ר; לערך פי 35 משטח העור החיצוני.
מן הדם שבנימים עובר אל חלל נאדיות הריאה – ומשם אל האוויר החיצוני בתהליך הנשיפה – פחמן דו-חמצני, CO2. פחמן דו-חמצני נוצר כתוצר לוואי של נשימת התאים. ואולם פחמן דו-חמצני נוצר לא רק בנשימה אאירובית, אלא גם בנשימה אלאווירנית (אנאירובית), שאינה עושה שימוש בחמצן מולקולרי.
מכל מקום, מַעבר החמצן המולקולרי מן האוויר לדם ומעבר הפחמן הדו-חמצני מן הדם לאוויר מכונה בפי כל "חילוף הגזים של הנשימה". אין בכך כל פסול. ובכל זאת, למען הדיוק נדמה שראוי להעיר בהקשר לכך שלושה דברים.
ראשית, מן התיאור "חילוף הגזים" ניתן אולי להסיק שקיים יחס נפחים של 1:1 בין החמצן שנקלט לבין הפחמן הדו-חמצני שנפלט מהדם לאוויר. הדבר מדויק כשמתרחש בתאים חימצון מלא של סוכרים באמצעות חמצן מולקולרי. אך היחס שבין O2 שנקלט ל-CO2 שנפלט אינו תמיד 1:1. למשל: חלק מהפחמן הדו-חמצני שנוצר בתאים של יונקים משמש ליצירת שתנן (אוריאה), המופרש כתוצר פירוק של חלבונים וחומצות גרעין. מכאן שחלק מה- CO2 כלל לא נפלט דרך הריאות, אלא מופרש בשתן כחלק ממולקולת השתנן.
ובאופן כללי יותר, כשתאי הגוף מפיקים אנרגיה כימית לא מפירוק של סוכרים כמו גליקוגן או גלוקוז, אלא מחומרים אורגניים אחרים, מספר מולקולות הפחמן הדו-חמצני הנוצרות בנשימת התאים נמוך ממספר מולקולות החמצן הנדרשות לנשימה. יחס זה CO2/O2, מכונה "מקדם הנשימה" (respiratory quotient).
בעוד שעבור חימצון סוכרים ערכו שווה כאמור 1, הרי שעבור שומנים ערכו נמוך יותר – מספר מולקולות הפחמן הדו-חמצני שנפלטות בעקבות חימצון נשימתי של שומן קטן ממספר מולקולות החמצן המולקולרי הדרושות לתהליך. כך גם לגבי חימצון נשימתי של חלבונים.
זימים במים
שנית, בעלי חיים רבים מאד – במקרה זה הביטוי "רבים מני ים" איננו רק ספרותי גרידא – סופגים חמצן מולקולרי לא מן האוויר, אלא מן המים. מדובר כמובן בחמצן מולקולרי המומס במי-הים, במי-האגם או במי-הנחל שבהם חיים בעלי-חיים, הסופגים חמצן דרך שטח הגוף החיצוני (מדוזות, למשל) או באמצעות איברי נשימה ספציפיים – זימים.

בניגוד לריאות (ולטרכיאות של חרקים), שהן שקעים פנימה לתוך הגוף, זימים הם בליטות כלפי חוץ (לעתים, אמנם, מוגנים ומכוסים). בליטות זקוקות בדרך-כלל לתמיכה, ואכן המדיום המֵימי מקנה תמיכה לזימים העדינים. זו אחת הסיבות לכך שנושמי זימים נחנקים באוויר, אף שתכולת החמצן בו גבוהה הרבה יותר מזו שבמים (ליטר אוויר בגובה פני הים מכיל לערך פי 30 מולקולות חמצן מולקולרי מאשר ליטר מים הרווים בחמצן מומס) – הזימים העדינים קורסים ונדבקים זה לזה אם אינם נתמכים על ידי כוח העילוי של מדיום מֵימי.
מבין החולייתנים, סופגים חמצן מולקולרי בעזרת זימים הדגים והכרישים, ראשני צפרדעים וקרפדות, וכן דו-חיים שאינם עוברים גלגול, דוגמת הפרוטיאוס האירופי והאקסולוטל המקסיקני. חסרי חוליות מימיים רבים נושמים גם הם בעזרת זימים: סרטנים, תולעים רבות, רבות מהרכיכות. והנה, בעלי חיים אלה מפיקים ATP תוך ניצול חמצן מולקולרי במיטוכונדריונים של תאיהם לשם זירחון חימצוני. אבל גזים של נשימה?! – רבים מהם יכולים לחיות את כל חייהם באושר מֵימי עילאי מבלי לחוש כלל חמצן ופחמן דו-חמצני כגזים! יתר על כן, חמצן גזי, חמצן מן האוויר, פירושו יכול להיות עבורם חנק (בין השאר, כאמור, עקב קריסת הזימים), כפי שכותב המשורר (והמדען) אבנר טריינין על דג שנלכד "[...] ים-אימים שנתייבש, כל כך הרבה / חמצן להחנק. לשווא כה יעלע בזימיו" (מתוך "משירי לאונרדו", שיר 2. מן הקובץ "השער הסתום", הקבה"מ, 1976).
 |
| אקסולוטל - דו-חי שמקורו במקסיקו, הנושם בעזרת זימים - לאמור, נושם חמצן מומס במים - גם בבגרותו (הזימים הם האיברים הבולטים לפני הרגליים הקדמיות). צילום: Amandasofiarana - Wikimedia commons |
זימים הם איברים "רב-שימושיים", ומעבירים לא רק את המומסים המשתתפים בתהליך הנשימה – O2 ו-CO2, אלא גם יונים המומסים במים, וזאת ללא קשר לנשימה. ואכן, דגי-ים מסלקים באמצעות הזימים עודפי מלחים (כגון Na+Cl-) שחדרו לגופם ממי הים, ואילו דגי נהרות ואגמים של מים דלי מלחים שואבים לגופם יונים מן הסביבה באמצעות זימיהם. וכמו שלא "עולה בדעתם" של נושמי הזימים שהמלחים הנשאבים אל (או מ-) גופם הם מוצקים כשאינם בצורה מומסת, כך לא "עולה בדעתם" שהחמצן והפחמן הדו-חמצני הם גזים – הם באים במגע עם רק עם תמיסות מימיות, לא עם גזים. לפחות כך הדבר כשמדובר בנשימה; לא כן כשמדובר בשלפוחית השחייה.
גז לריחוף, לא לנשימה
בגופם של דגים רבים יש שלפוחית שחייה, המאפשרת לדג לרחף בגוף המים מבלי להשקיע אנרגיה בניסיון לא לשקוע לקרקעית, למרות שהמשקל הסגולי הממוצע של רקמות חיות עולה על זה של המים – של מי-ים ובוודאי של מים דלֵי מלחים. שלפוחית השחייה מגדילה את הנפח הכולל של גוף הדג (לערך ב-5% בדגי-ים וכ-7% בדגי מים מתוקים) מבלי להוסיף כמעט משקל, וכך קטֵן המשקל הסגולי הכולל של הדג, עד שהוא הופך "נייטרלי" – שווה בדיוק לזה של המדיום, מה שמאפשר לדג לרחף במים ללא מאמץ.
שלפוחית השחייה של דגים נוצרת כשקע של צינור העיכול; בחלק מן הדגים צינור ממשיך לחבר את השלפוחית אל הלוע, אך בדגים רבים מאד, בשלב מסוים בהתפתחות ניתק הקשר, וחלל השלפוחית הופך לחלל סגור, מנותק מן העולם החיצוני. בדגים אלה הרכב הגזים שבשלפוחית השחייה יכול להיות שונה מאד מזה שבאוויר החיצוני.
יש דגים ששלפוחית השחייה שלהם מכילה חנקן כמעט טהור, ומעט גזים אצילים; בדגים אחרים הגז העיקרי בשלפוחית השחייה הוא חמצן, O2. בשני המקרים ברור שאין מדובר ב"גזים של הנשימה", שהרי החנקן והגזים האצילים אינרטיים לחלוטין מבחינת תהליך הנשימה, ואילו החמצן נדרש כמובן לנשימה, ולא מופרש בה. מכאן שנושמי זימים שלהם שלפוחית שחייה, מצב צבירה גזי קשור אצלם בבקרת המשקל הסגולי ובריחוף במים, אך לא לנשימה.
ושלישית, כשם שנושמי הזימים אינם מכירים חמצן מולקולרי בצורת גז בהקשר לנשימה אלא כחלק מהתמיסות המימיות – התמיסות שבתוך גופם, וזו שמהווה את המדיום בו הם חיים, כך גם המיטוכונדריונים, צרכני החמצן שבגופנו: גם הם עוסקים בחמצן ובפחמן דו-חמצני רק כשהם חלק מתמיסה מימית, לא כפאזה גזית.
החמצן המולקולרי הנמצא בחלל נאדיות הריאה בפאזה גזית עובר לנימי הדם, ונקשר ברובו המכריע להמוגלובין שבתאי הדם האדומים וכך מובל בזרם הדם; מעט החמצן מובל כשהוא מומס, כחלק מהפאזה הנוזלית – פלזמת הדם. ברקמות הגוף ההמוגלובין משחרר חמצן מולקולרי הנותר תמיד בפאזה נוזלית – פלזמת הדם, נוזל הרקמות, ציטופלזמת התאים והתמיסה המימית שבתוך המיטוכונדריונים.
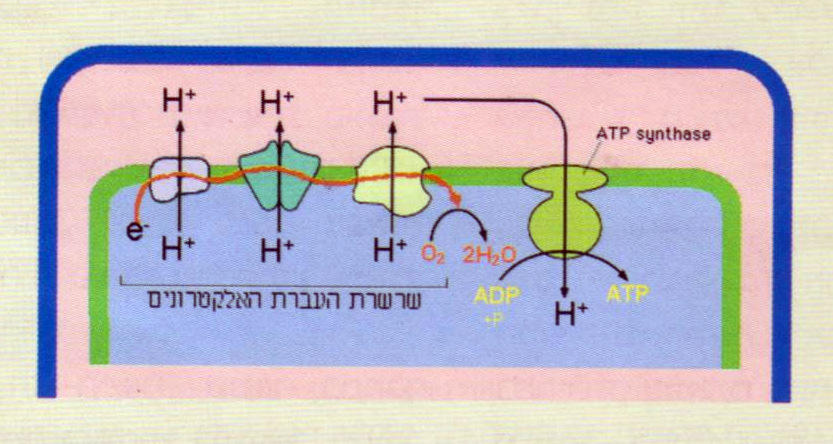
בתמיסה שבמיטוכונדריונים מקבל החמצן אלקטרונים (שמקורם בחומר האורגני המתחמצן בנשימה) מ"שרשרת העברת האלקטרונים", ונקשר לפרוטונים (H+). לפיכך החמצן, שהיה מומס בפאזה המימית, יוצר בסופו של תהליך, מים, מֵי-נשימה.
הפחמן הדו-חמצני, שנוצר במהלך פירוק החומר האורגני בנשימה, מועבר בדם ברובו בצורת יון ביקרבונט (HCO3-) מסיס במים; מעט ממנו עובר בצורת CO2 מומס בפאזה המימית – פלזמת הדם, בחלקו מועבר כשהוא קשור לחלבונים, וחלקו נכלל, כזכור, במבנה השתנן המובל בדם ומופרש בשתן. מכל מקום, הפחמן הדו-חמצני מופיע כגז רק בנאדיות הריאה; לא בתאי הגוף ולא בדם.
סכנה – בועות גז
הופעה של גז – גז ממש, בצורת בועות – בתוך כלֵי הדם, היא תופעה מסוכנת, המתבטאת במחלת הדֶקומפרֶסיה. מחלת הדקומפרסיה נגרמת עקב נשימת אוויר (או תערובת גזים אחרת) בלחץ גבוה יחסית, ומַעבר מהיר מדי לנשימת אוויר בלחץ נמוך יותר, כמו למשל בעת עלייה מהירה מדי מצלילה. על-פי חוק הֶנרי, ככל שלחצו של גז גבוה יותר, גדלה מסיסותו בנוזל אתו הוא בא במגע. לפיכך בעת צלילה מתמוססים יותר גזים בפלזמת הדם וכן בנוזלי גוף אחרים, כגון בנוזל המפרקים. והנה, אם הלחץ יורד במהירות, הנוזלים נעשיים רוויים בגזים, ובעיקר בחנקן. עודף הגז יוצא מן התמיסה, כלומר מן הפאזה המימית, ויוצר פאזה גזית – בועות בכלֵי הדם ובנוזל המפרקים. אלו גורמות כאבים קשים במפרקים, ועלולות לחסום כלי דם ולמנוע הספקת דם לאזורים חיוניים. מסיבה זו מחלת הדקומפרסיה עלולה לפגוע במוח השִדרה ולגרום לשיתוק פלג הגוף התחתון (פּראפְּלֶגיה), או אף לשיתוק נרחב יותר ולפגיעות במוחות הגולגולת.
בהקשר זה מעניין לשוב ולבחון את שלפוחית השחייה בדגים: כשדג צולל לעומק רב יותר, הלחץ על גופו עולה, כתוצאה מכך הלחץ בתוך גופו עולה, כולל בשלפוחית השחייה שלו. אם הלחץ גדל, הרי שלפי חוק בּוֹיְל נפח השלפוחית חייב לקטון (חוק בויל: כששאר התנאים קבועים, מכפלת לחץ הגז בנפחו היא גודל קבוע). אבל אם נפח השלפוחית קטֵן, קטן הנפח הכללי של גופו, משקלו הסגולי גדל והוא עלול לצלול כאבן למעמקים (או להשקיע מאמץ שרירים בשחייה מאומצת כלפי מעלה). יתר על כן, כשהלחץ גדל, גז משלפוחית השחייה נוטה (לפי חוק הנרי) להתמוסס בנוזלי הגוף, ולפיכך נפח השלפוחית יקטן עוד יותר, ובעיית הדג עוד תכבד, פשוטו כמשמעו.
והנה, מתברר כי דגים בעלי שלפוחית שחייה מצוידים במנגנונים אנטומיים ופיזיולוגיים מופלאים הדואגים לכך שבעת צלילה מאסת הגז בשלפוחית השחייה לא רק שאיננה קטנה (עקב התמוססות), אלא אף גדלה, כך שנפח השלפוחית נותר בעינו, והדג ממשיך לרחף במים – משקלו הסגולי הכללי נותר שווה לזה של המים בהם הוא שוחה. ממש דג במים.
* * *
לסיכום: לרבים-רבים מבעלי החיים דרוש החמצן "כמו אוויר לנשימה" לצורך מיצוי אנרגיה מחומרים אורגניים בתהליך החימצון במיטוכונדריונים ובניית ATP. תהליך זה תלוי בתכונותיו הכימיות של החמצן – זיקתו הגבוהה לאלקטרונים, ולא במצב הצבירה שלו. המיטוכונדריונים העושים את השימוש בחמצן כלל אינם נחשפים לחמצן כגז, ורבים מנושמי חמצן מומס באמצעות זימים (או שטח הגוף) כלל אינם באים במגע עם גזים, לפחות לא בהקשר של תהליכי הנשימה.
גליקוליזה, מעגל קרבס, שרשרת העברת האלקטרונים והמנגנון הכימואוסמוטי הסוכר גלוקוז - מולקולה 6 פחמנית ( C6) שמגיעה לתאים מהדם, או נוצרת (בעיקר בתאי שריר) מגליקוגן, רב־גלוקוז המשמש כחומר תשמורת - עובר בציטופלזמת התא סדרה של שינויים, שבסופה נוצרות 2 מולקולות תלת-פחמניות של פירובט (C3). סדרת תהליכים אנזימטיים זו מכונה גליקוליזה, ובמהלכה נוצר ATP. הפירובט חודר לחלל הפנימי של המיטוכונדריונים, שם פועל עליו מערך אנזימים המכונה פירובט-דהידרוגנז (וראו: ״תיאמין לא אמין״, גליליאו 64). בעקבות הפעולה משתחרר CO2 והפירובט הופך לאצטיל C2. האצטיל, הקשור למולקולה מורכבת המכונה קואנזים A, (ובקיצור קו-A). מגיב עם חומר ארבע-פחמני C4 תוך יצירת חומצת לימון (C6). חומצת הלימון עוברת סדרה של שינויים כימיים בעזרת אנזימים, במהלכה משתחררות שתי מולקולות CO2 וכך שב ומופיע החומר הארבע-פחמני שמסוגל לשוב ולקשור קבוצת אצטיל, וחוזר חלילה - זהו, אם כן, מעגל סגור, ״מעגל קרבס״ (המכונה גם מעגל חומצת הלימון). בחלק מן התהליכים הכלולים במעגל קרבס מופקת אנרגיה בכמות המאפשרת בניית ATP, ואולם האנרגיה הכימית שהיתה כלולה במערכת המקורית גלוקוז/חמצן נותרה עדיין ברובה - בשלב זה היא אגורה בחומרים מחזרים, שמכילים את המימן שמקורו בגלוקוז (כל אטומי הפחמן של הגלוקוז כבר כלולים בשלב זה בחומר אי-אורגני דל אנרגיה, פחמן דו-חמצני).
חומרים מחזרים אלה מוסרים את האלקטרונים, שמקורם באטומי המימן שנותרו מהגלוקוז, לשורה של חומרים - שרשרת העברת האלקטרונים, שרכיביה שוכנים בקרומית הפנימית של המיטוכונדריון, בעוד הפרוטונים (אטום מימן בנוי מפרוטון כגרעין ואלקטרון המקיף אותו) נותרים בתמיסה הפנימית. בסופו של דבר נמסרים האלקטרונים לחמצן המולקולרי הנמצא בתמיסר המימית, ובכך מגיעים האלקטרונים ל״מנוחה נכונה׳׳. האנרגיה שלהם אז נמוכה מאוד. החמצן שזכה באלקטרונים נקשר לפרוטונים וכך נוצרות מולקולות מים; אלה הם מי-נשימה. במהלך מעבר האלקטרונים מרכיב לרכיב בשרשרת העברת האלקטרונים בדרכם אל החמצן, האנרגיה שלהם פוחתת והולכת! חלק מהאנרגיה נרתם על ידי רכיבי השרשרת לשאיבת פרוטונים מן החלל הפנימי אל חלל הביניים. כך נוצרים בחלל הביניים ריכוז גבוה של פרוטונים ופוטנציאל חשמלי חיובי (שהרי הפרוטונים הם יונים חיוביים). פירוש הדבר הוא שלפרוטונים בחלל הביניים אנרגיה גבוהה והם נדחקים לחזור לחלל הפנימי. ואולם הקרומית הפנימית אינה מאפשרת להם לחזור, והדרך היחידה הפתוחה בפניהם היא מעבר מבעד לאנזים ATP-סינטז, המנצל את אנרגיית הפרוטונים הנדחקים לחזור לבניית ATP, מ-ADP וקבוצה זרחתית. זהו המנגנון הכימואוסמוטי: יצירת הפרש של ריכוזי פרוטונים, ורתימת הפרש זה לבניית ATP. לא רק פירובט שמקורו בגלוקוז יכול ”לתדלק׳׳ את מעגל קרבס ואת שרשרת העברת האלקטרונים ובעקבותיה את המנגנון הכימואוסמוטי ותהליך בניית ה-ATP. יכולות לעשות זאת גם חומצות אמינו שעברו דאמינציה (ניתוק המרכיב החנקני מהמולקולה) - חלקן הופכות לפירובט, חומצות אמינו אחרות הופכות לאצטיל, ויש חומצות אמינו המשתלבות בשלבים שונים של מעגל קרבס עצמו. חומצות שומן ׳׳מתדלקות״ יצירת ATP דרך אצטיל קו-A. מכל מקום, אטומי מימן (בצורת פרוטונים שבתמיסה, ובנפרד אלקטרונים המובלים בשרשרת העברת האלקטרונים) שמקורם בחומרים האורגניים, נקשרים לחמצן מולקולרי שהוא חלק מן התמיסה המימית שבחלל הפנימי במיטוכונדריונים, וכך נוצרים מים המתוספים ישירות לתמיסה. בכל השלבים הללו מולקולות החמצן אינן מופיעות בפאזה גזית אלא בתמיסות מימיות!
|

פורסם ב"גליליאו" 70, יוני 2004